
Vol. 4, N° 12. Setiembre 2017 pp. 46-68
Artículos
La vid silvestre. Un importante recurso fitogenético sin protección legal en España The wild vine.
An important phytogenetic resource without legal protection in Spain
Lara, M. Iriarte-Chiapusso, M.J., Cantos, M., García Jiménez, J.L., Morales, R., Ocete, C.A., López, M.A., Salinas, J.A., Rubio, I., Hidalgo, J., Íñiguez, M., Rodríguez, A., Valle, J.M., Arroyo-García, R., Ayala, M.C., Armendáriz, I., Maghradze, D., Arnold, C. y Ocete, R.
Lara, M.: IFAPA, Rancho de la Merced, Jerez de la Frontera (Cádiz). Correo electrónico: miguel.lara@juntadeandalucia.es.
Iriarte-Chiapusso, M.J.: Área de Prehistoria, IKERBASQUE/Universidad del País Vasco (Vitoria/Gasteiz).
Cantos, M. y García Jiménez, J.L.: Grupo de Investigación Propagación y Nutrición Vegetal, IRNAS-CSIC (Sevilla). Correo electrónico: cantos@irnase.csic.es.
Morales, R.: Real Jardín Botánico-CSIC (Madrid). Correo electrónico: morales@rjb.csic.es.
Ocete, C.A. y López, M.A.: Laboratorio de Entomología Aplicada, Facultad de Biología, Universidad de Sevilla. Correo electrónico: ocete@us.es.
Salinas, J.A.: Jardín Botánico de Miranda de Ebro (Burgos).
Rubio, I.: Herbario de los Montes Obarenes, Miranda de Ebro (Burgos).
Hidalgo, J.: Aunia Kultura Elkartea, Laudio (Vizcaya). Correo electrónico: juanjoaunia@gmail.com.
Íñiguez, M.: Estación Enológica de Haro (La Rioja).
Rodríguez, A. y Valle, J.M.: Escuela Universitaria de Ingeniería, Universidad del País Vasco (Vitoria/Gasteiz).
Arroyo-García, R.: Centro de Biotecnología y Genómica de Plantas, INIA, Pozuelo de Alarcón (Madrid).
Ayala, M.C.: Instituto La Laboral (Lardero, La Rioja).
Armendáriz, I.: Universidad Tecnológica Equinoccial (Quito, Ecuador).
Maghradze, D.: Institute for Horticulture, Viticulture and Oenology (Tbilisi, República de Georgia).
Arnold, C.: Universidad de Lausana (Suiza)
Ocete, R.: Laboratorio de Entomología Aplicada, Facultad de Biología.
Universidad de Sevilla. Correo electrónico: ocete@us.es.
Resumen
El presente artículo está enfocado a la necesidad de conseguir en España una figura de protección legal del recurso fitogenético vid silvestre euroasiática, el parental dioico de las variedades de cultivo. Tras señalar los distintos tipos de hábitats que ocupa en la Península Ibérica, se indican las principales características ampelográficas de las plantas y su estado sanitario. Por otra parte, se enumeran los usos antiguos que el ser humano ha hecho de las mismas y se analizan los impactos antrópicos negativos que han llevado a considerarlas un taxón amenazado, señalando los distintos tipos de hábitats que ocupa en la Península Ibérica. Resaltan las características más importantes de las vides silvestres para llevar a cabo programas de mejora genética, tanto de portainjertos como de viníferas, para abordar las consecuencias de la erosión genética del viñedo en el actual marco de cambio climático. Se recogen además las medidas de conservación que se han puesto en marcha hasta ahora en distintas zonas de España, para finalmente adjuntar un listado de centros en donde se investigan distintas facetas de este legado silvestre.
Palabras clave: conservación de germoplasma, mejora genética, taxón amenazado, usos tradicionales, Vitis vinífera subsp. sylvestris.
Abstract
The present article is focused on the need to get in Spain a legal figure of legal preservation for the phytogenetic resource Eurasian wild grapevine, the dioecious parental of the cultivated varieties. It shows its different habitats in the Iberian Peninsula. The main ampelographical characteristics of these grapevines and their sanitary status are included. On the other hand, it enumerates the ancient uses that the man has made of these grapevines and the negative anthropic impacts which have made them a threatened taxon. The most important characteristics of the wild grapevines are remarked, in order to carry out genetic breeding programs of rootstocks and cultivars to reduce the consequences of genetic erosion of the vineyards in the present context of climate change. It includes some conservation measures that have been implemented until now in several Spanish regions. Finally, a list of research centers with different lines of investigation on this wild legacy is also attached.
Keywords: breeding, measures of preservation, traditional uses, threatened taxon, Vitis vinífera subsp. sylvestris
Introducción
El taxón Vitis vinifera L. subsp. sylvestris (C.C. Gmelin) Beger & Hegi constituye la única especie de vid autóctona de Eurasia. Virgilio le dio el nombre de labrusca, que se sigue conservando en las lenguas romances. Sus poblaciones se extienden por diversos ecosistemas naturales desde la Península Ibérica hasta Afganistán, concretamente hasta el macizo del Hindu Kush (Arnold, 2002). También en la zona de El Maghreb pueden encontrarse poblaciones de esta vitácea, como es el caso del río Ourika, al pie de la cordillera del Atlas, al sur de Marruecos (Ocete et al, 2007).
Las parras silvestres son lianas hidrófilas y heliófilas; por este último motivo, estas lianas se encuentran dotadas de zarcillos para poder trepar por ejemplares de porte arbóreo o arbustivo de la vegetación circundante (Figuras 1 y 2). Esta característica evolutiva podría deberse a las cálidas condiciones climáticas del Paleoceno, que propiciaron la expansión de densos bosques en los que las vitáceas y otras familias botánicas tuvieron que verse forzadas a competir por la luz, para obtener una intensidad lumínica adecuada para su proceso fotosintético (Mai, 1987). La vid silvestre euroasiática constituye el parental dioico de las variedades de cultivo, englobadas en Vitis vinifera L. subsp. sativa DC. ex Hegi, integrada mayoritariamente por cultivares hermafroditas seleccionados por el hombre (Arnold, 2002).
Parece ser que la Península Ibérica constituyó uno de los refugios europeos para la vid silvestre durante el Pleistoceno. De hecho, entre otros, existen acúmulos de polen en los niveles D2 de Cal Guardiola (Terrasa) (Postigo et al, 2007) y en el TD7 de la Gran Dolina de la Sierra de Atapuerca (García-Antón, 1989). Estos últimos hallazgos, con una antigüedad de unos 780.000 años, se sitúan justo en el límite entre el Pleistoceno inferior y medio (Discontinuidad de Brunhes-Matuyama), época en la que se produjo la última inversión de los polos magnéticos.
Figuras 1 y 2. Vides silvestres trepando por la vegetación circundante
Fuente: archivo personal.
Al terminar el último periodo glaciar (Würm), las nuevas condiciones ambientales y la expansión de los bosques mesotermófilos facilitaron la recuperación de la vid silvestre y otras especies botánicas en dicho territorio (Iriarte-Chiapusso et al, 2005). Dentro del País Vasco, aparecen granos de polen de esta vitácea en la zona de Urdaibai (Vizcaya) correspondientes a los comienzos del Holoceno, concretamente del décimo milenio a.C. (Iriarte-Chiapusso et al, 2006).
En Eurasia, la máxima diversidad de variedades de vid se da en la zona del Sur del Cáucaso, por lo que el gran investigador soviético Vavilov (1926) supuso que era la zona geográfica en la que se había producido el primer fenómeno de domesticación de las parras silvestres. Más tarde, Negrul (1938) bautizó a esa zona con el nombre de Triángulo de la Uva Fértil de Vavilov. Curiosamente, el Monte Ararat, donde la tradición supone que encalló el Arca de Noé, y donde este se embriagó, se encuentra dentro de dicha área geográfica.
Con estos antecedentes, los principales objetivos de este artículo son concentrar en una publicación holística los conocimientos sobre la vid silvestre en nuestro país, con el fin de intentar lograr una figura legal para su protección.
Hábitats
Las actuales poblaciones relictas de vid silvestre, dentro de España, corresponden a manchas mucho más pobladas que se conservaron hasta el siglo XIX. En efecto, la llegada de las enfermedades fúngicas norteamericanas oídio y mildiu supusieron un fuerte impacto sanitario sobre las vides europeas tanto cultivadas como silvestres (Ocete et al, 2004). Los ejemplares silvestres se distribuyen fundamentalmente en núcleos poblacionales dispersos, incluidos en bosques de ribera, formaciones azonales de las orillas de ríos y arroyos de todo el territorio nacional (Figura 3). En las zonas con mayor precipitación también pueden encontrarse en posición coluvial, como es el caso del norte de León, Asturias, Cantabria y País Vasco. En áreas litorales se distribuyen por los acantilados y playas del Cantábrico, desde el País Vasco, incluyendo el territorio de Lapurdi (Iparralde), hasta el poniente de Asturias (Figura 4). Otras poblaciones se encuentran en arenosoles de las proximidades a la desembocadura del Guadalquivir, en la Algaida (Sanlúcar de Barrameda) y en el Parque Nacional de Doñana (Ocete et al, 1999, 2007).
Figura 3. Foz de Arbayún (Navarra)

Fuente: archivo personal.
Figura 4. Población del litoral Cantábrico (Asturias)

Fuente: archivo personal.
Principales características ampelográficas y origen de las variedades de cultivo
En el mapa a continuación apreciamos aquellas provincias donde hemos encontrado poblaciones de vides silvestres (Figura 5).
Figura 5. Mapa con indicación de las provincias donde se han encontrado poblaciones de vides silvestres
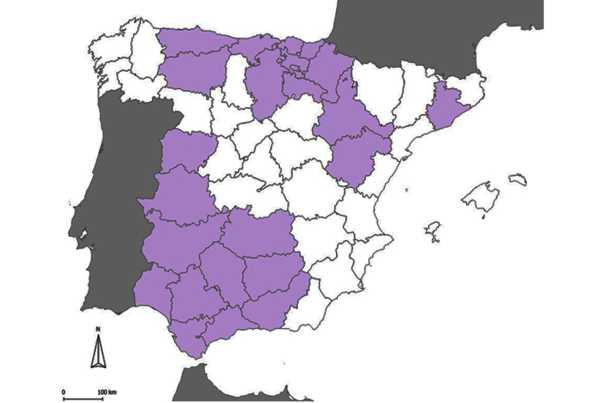
Fuente: elaboración propia.
Las plantas masculinas se caracterizan por tener gran cantidad de racimos con flores tipo I, integradas por cinco estambres y ausencia de gineceo. No obstante, ocasionalmente, aparecen ejemplares con flor de tipo II, con un ligero desarrollo de gineceo, según la nomenclatura propuesta por OIV (2009). En estas plantas, a veces, llega a formarse un escaso número de bayas por racimo.
El polen masculino es tricolporado, semejante a los ejemplares hermafroditas de cultivo, con orificios para la salida del tubo polínico por el que saldrán los gametos. Las flores femeninas presentan un gineceo bien desarrollado, estambres reflejos y polen acolporado, carente de orificios para la salida del tubo polínico (Gallardo et al, 2009) (Figuras 6 y 7). En ambos sexos el ápice superior de los brotes es abierto. Generalmente, el tamaño de las hojas de las plantas masculinas, que suelen ser más lobuladas, es inferior al de las femeninas. Comúnmente el seno peciolar es muy abierto en las primeras y abierto en las segundas (Benito et al, 2016).
Figura 6. Flores masculinas, tipo I y II;
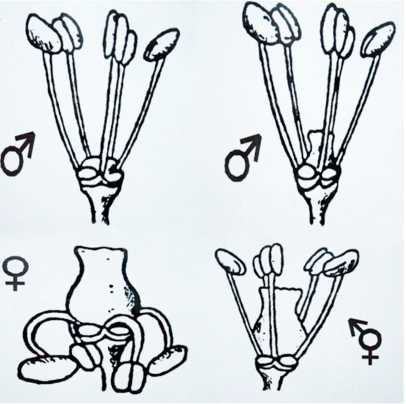
Fuente: archivo personal.
Figura 7. Ejemplar masculino en plena floración femenina y hermafrodita

Fuente: archivo personal.
Las parras femeninas producen racimos con bayas que no superan el centímetro de diámetro. La inmensa mayoría son tintas, con hollejo azul oscuro y pulpa no coloreada (Figura 8). Las pepitas son más redondeadas, con la chalaza más corta que las procedentes de variedades de cultivo, como puede apreciarse en la Figura 9. Este hecho permite saber si en un determinado yacimiento arqueológico se consumían bayas silvestres o de cultivo.
Figura 8. Racimo en fase de maduración

Fuente: archivo personal.
Figura 9. Diferencias morfológicas entre semillas cultivadas (parte superior) y silvestres
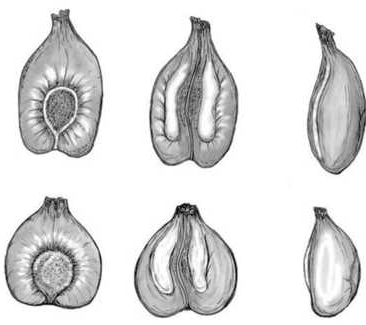
Fuente: archivo personal.
De acuerdo con lo expuesto en los párrafos anteriores, la mutación de ejemplares masculinos daría lugar a plantas hermafroditas que tendrían un mayor cuajado de frutos. Estas fueron seleccionadas por el hombre y sus sarmientos plantados fuera de los hábitats naturales, iniciándose así su cultivo. El refugio de Transcaucasia, protegido de los vientos de componente norte por las elevadas cumbres de la cordillera del Cáucaso, sirvió de albergue a varias especies de frutales, incluidas las parras silvestres, durante las glaciaciones del Pleistoceno (McGovern, 2004).
Las semillas de vid cultivada más antiguas, encontradas hasta el momento, tienen una antigüedad de unos 8.000 años a.C. Proceden del tell del yacimiento arqueológico de Shulaveri Gora (República de Georgia), dentro del citado Triángulo de Vavilov (Chlilashvili, 2004) (Figuras 10 y 11).
Figura 10. Semillas de tipología cultivada del yacimiento arqueológico de Shulaveri Gora (Georgia), hacia el 8.000 BP

Fuente: Museo de Tbilisi.
Figura 11. Recipiente cerámico que ha contenido mosto o vino, según el análisis de sus residuos (Shulaveri Gora, Georgia)

Fuente: Museo de Tbilisi.
Incidencia de plagas y enfermedades
En este apartado, haremos referencia a la zona radicular y a los órganos aéreos.
Referente a los pelos absorbentes radiculares, cabe destacar la ausencia de síntomas atribuibles a la fase radicícola de la filoxera, Daktylosphaera vitifoliae (Fitch), así como de nódulos provocados por nematodos (Ocete et al, 2011 a y b). Cabe señalar que tampoco se han observado micelios blanquecinos de hongos asociados a la podredumbre causada por Armillaria mellea (Vahl.) Kumm., los cuales, sin embargo, son bastante frecuentes en los chopos que, en mucha riberas, acompañan a las parras, sirviéndoles incluso de tutor.
La ausencia de infestación por filoxera y de nódulos causados por nematodos parece deberse a las condiciones especiales del perfil de los fluvisoles sobre los que se asientan la mayor parte de estas poblaciones, que sufren dilatados periodos de encharcamiento cada año.
No obstante, fuera de su medio natural las raíces de nuestras vides silvestres sí son sensibles a la fase radicícola de la filoxera (Ocete et al, 2011a). En relación con estos hechos, en Francia, a comienzos de la reconstrucción postfiloxérica, Camile Saint Pierre señalaba que los sarmientos de los ejemplares silvestres se llegaron a emplear como portainjerto en algunos viñedos reconstituidos. Al haberlos sacado y plantado fuera de su ambiente natural, empezaron a aparecer síntomas de infestación con el consiguiente deterioro de la apariencia de las cepas y su muerte (De La Blanchére, 1876).
Sobre las hojas son frecuentes las falsas agallas causadas por el ácaro de la erinosis, Colomerus vitis (Pagenstecher) (Acari, Eriophyidae). En menor proporción se han observado síntomas de acariosis, causados por Calepitrimerus vitis (Nalepa) (Acari, Eriophyidae). En cuanto a los síntomas causados por oídio, Erysiphe necator (Schwein.) Burriel, Oidium tuckeri (Berck) y mildiu, Plasmopara viticola (Berlease & de Toni), son palpables tanto en los órganos vegetativos como en los racimos, desde su cuajado hasta el estado de maduración de las bayas, a finales de octubre o comienzos de noviembre.
Los niveles de infestación y de infección varían de un ejemplar a otro dentro de cada núcleo poblacional, dada la diversidad genética existente entre los ejemplares de una misma población.
Lo expuesto anteriormente indica que ambos ácaros eriófidos monófagos pasaron de los ejemplares silvestres a los cultivados durante el proceso de domesticación de la vid. En sentido opuesto, los citados hongos, importados de Norteamérica durante el siglo XIX sobre ejemplares de vid americana, han pasado del viñedo a las poblaciones silvestres de la zona (Ocete et al, 2011a; Maghradze et al, 2015).
Por otra parte, cabe resaltar que los diversos Test ELISA para la determinación de virosis siempre han dado negativo (Arroyo-García et al, 2006).
Usos antiguos de la vid silvestre
Seguidamente, se recogen algunos de los usos que las vides silvestres han tenido para el hombre desde el Pleistoceno superior hasta la actualidad. Todos ellos son tomados de fuentes bibliográficas, que naturalmente se citan en cada caso.
Uso como combustible de la madera y de las bayas como alimento
En el caso del yacimiento arqueológico del Abric Romaní (Anoia, Barcelona), el análisis de la madera quemada ha revelado restos de lianas de vid, entre otros restos vegetales, que fueron empleados por los neandertales como combustible (Burjachs y Juliá, 1994).
Los restos arqueobotánicos procedentes de yacimientos arqueológicos de la Península Ibérica evidencian que la vid silvestre ha sido un recurso vegetal empleado por el ser humano desde el Paleolítico, formando parte de su dieta alimentaria (Rivera y Walker, 1989). A lo largo del Holoceno esta planta se extiende por el territorio peninsular, siendo frecuente en los ecosistemas mediterráneos. Al inicio de este periodo, las últimas sociedades cazadoras-recolectoras del Mesolítico mantendrán su consumo (Guerrero, 1995) al igual que las sociedades productoras que les suceden (Neolítico, Calcolítico, Edad del Bronce), como lo atestiguan los registros carpológicos de yacimientos como La Draga (Banyoles), Can Tintorer (Barcelona), Los Millares (Almería), Cova Farisa (Huesca) o Castellón Alto (Granada) (Alonso, 1995; Rodríguez-Ariza et al, 1996). A partir de la Edad del Hierro, la vid cultivada adquirirá mayor protagonismo.
Usos enológicos y fabricación de vinagre
En la etapa previa a la domesticación de la vid, los racimos de bayas silvestres femeninas constituyeron la materia prima del vino. Carbonell (1820) señalaba lo siguiente:
Hay pocas producciones naturales que el hombre se haya apropiado para su sustento sin alterarlas, ó modificarlas, por medio de preparaciones, que varian mucho de su estado primitivo: la harina, la carne, los frutos, todo recibe de la mano del hombre un principio de fermentación antes de servirle de alimento, hasta los objetos de puro lujo, de capricho, ó de imaginacion, como el tabaco, los perfumes... Deben al arte peculiares modificaciones: pero la fabricación de los licores es en donde el hombre ha desplegado todo su ingenio, pues exceptuando el agua y la leche, todos los demas son obra suya. La naturaleza jamás formó licores espirituosos, lo que ella hace es promover la putrefaccion del racimo en la cepa, mientras que su jugo se convierte por el arte en un licor agradable, tónico y nutritivo, que llamamos vino.
Es difícil señalar la época en que los hombres comenzaron á fabricarle. Este precioso descubrimiento parece que se pierde en la oscuridad de los tiempos; y asi la invencion del vino tiene sus fábulas, del mismo modo, que la de todos los objetos, cuya utilidad ha llegado á ser general.
Al respecto, Quer y Gómez Ortega (1784: 479) señalaban lo siguiente: “Las Uvas de la Vid silvestre son estípicas, y suelen mezclarse sus racimos y flores en el mosto para comunicar al Vino mayor duración, y un cierto sabor raspante, agradable al gusto, y confortativo del estómago”.
En muchos casos, a los vinos blancos del año anterior se les añadían racimos de flores masculinas, con el fin de conferirles un aroma más afrutado. Dicha práctica ha sido muy común en otras áreas vitivinícolas, como es el caso de Azerbaijan, según comunicación personal de los doctores Salimov y Musayev.
En Alemania y Austria, la producción de vino con bayas silvestres se ha mantenido hasta épocas recientes. Además, en ciertas regiones de Italia y Alemania, los racimos silvestres se mezclaban con los cultivados para abaratar la producción de vinos caseros (Schumann, 1971; Anzani et al, 1993). En la isla de Cerdeña, un códice que contiene una serie de leyes de la segunda mitad del siglo XIV, denominado Carta de Logu, castiga la venta de vid silvestre, ya que no pagaba los impuestos ni diezmos y primicias. Con sus bayas se elaboraban los denominados vinu de marxani o vino de caprai (Lovicu et al, 2009). En dicha isla hemos recogido testimonios directos sobre la producción de vino de vulpa por los pastores, con la misma materia prima.
Hasta hace unos 20 años hemos observado la recogida de racimos de uva en los alrededores de la Serranía de Grazalema (Cádiz) para la fabricación casera de vinagre. No hay que olvidar que el vinagre y la sal fueron los mejores conservantes de alimentos en el pasado.
Usos medicinales
En la obra de Font Quer (1999), se recogen varios usos medicinales de las vides silvestres con sus oportunas referencias bibliográficas. Así, sobre la savia que brota durante la etapa de lloro, Quer y Gómez Ortega (1784: 479) refirieron: “De la Vid, así silvestre, como de la cultivada, se hacen varios usos en la Medicina. El Agua que naturalmente suelta ó llora en la Primavera, es aperitiva, diurética y buena para los ojos, para cuyas enfermedades la conservan algunos”.
Laguna (1555: 505-506) en referencia a la vid silvestre, señalaba:
La cual, no obstante que nos debe muy poco por haber sido de nosotros ansi olvidada, todavía se esfuerza de servirnos y complacemos con cuanto puede, quiero decir con su fructo, con sus tallos, con sus hojas y con sus flores en infinitas necesidades, principalmente cuando cumple y restriñir notablemente algún miembro, las cuales facultades se hallan mucho más eficaces en la labrusca, y principalmente en aquella que produce la enanthe, que en la vid cultivada...
Respecto al uso de racimos sin finalizar su maduración, estos han sido utilizados para fabricar un refresco, denominado agraz. Sobre las propiedades medicinales del mismo, Quer y Gómez Ortega (1784: 479) señalaban: “Las Uvas inmaduras o Agraces, y su zumo sirven para refrescar y restriñir, excitan el apetito, reprimen el ardor de las calenturas, y contienen las cámaras”. Ya Bustamante (1971) recoge el empleo del jugo de los racimos para remediar molestias estomacales en la zona más septentrional de Burgos, contenido en la Noticia Histórico Corográfica del Muy Noble y Real Valle de Mena, fechada en 1796: “Hállanse muchas parras en los montes y en los costados de los caminos y ríos y su fruto es muy bueno para agua de agraz”.
Usos en rituales funerarios
A lo largo de diversas culturas asentadas en España, los racimos de uva han sido empleados como ofrenda en rituales funerarios. Por ejemplo, en el caso de Andalucía han aparecido numerosos enterramientos con pepitas de vid silvestre en yacimientos de Córdoba (Torreparedones, Cerro de la Cruz-Almedinilla): Bronce Final/Hierro-Ibérico Pleno (Jones y Reed, 2000; Arnanz, 2000); Almería (Las Pilas-Mojácar; Los Millares): Calcolítico (Rovira Buendía, 2000); Granada (Castellón Alto-Galera, Cuesta del Negro): Bronce (Buxó, 1997); o Cádiz (Castillo de Doña Blanca): Bronce Final/Hierro (Chamorro, 1991).
En épocas más cercanas, se tiene constancia de enterramientos romanos donde han aparecido vasijas con semillas de vid cultivada. Respecto a este uso, ya se hacía eco Ovidio (s. I), con estas palabras:
Es un testimonio de respeto el tratar de aplacar en sus sepulturas a las almas de los antepasados y llevarles allí modestas ofrendas, pues los Manes exigen pequeñas cosas: coronas votivas, unas semillas de vid, unos pocos granos de sal, dones de Ceres empapados en vino y algunas violetas.
De acuerdo con las investigaciones llevadas a cabo por Torres-Vila y Mosquera-Müller (2001), en la zona de Los Bodegones (Mérida) -un importante complejo funerario romano-paleocristiano de los siglos IV-V- aparecieron semillas silvestres mezcladas con cultivadas. Los autores suponen que se llevó a cabo la plantación de un jardín funerario donde se desarrollaron cepas de ambos tipos de vid. Esos racimos mitificaban símbolos como el vino eucarístico, la resurrección y la comunión con ágapes funerarios.
Otros usos
Quer y Gómez Ortega (1784: 480) refieren: “Los mismos sarmientos de las Parras silvestres, o no podadas, se tuercen para formar maromas, que sirven de amarras para las barcas, para las redes de pescar, y para otros muchos objetos”.
A lo largo de 30 años de investigación sobre la vid silvestre, hemos visto que las plantas del Parque Natural de los Alcornocales (Cádiz) eran empleadas hasta hace unos 20 años para fabricar aros de las nasas de pesca empleadas por la flota de Barbate (Ocete et al, 2007). El mismo uso se les daba también en zonas del litoral asturiano.
Por otra parte, el ácido tartárico presente en el mosto ha servido como mejorante de pastas cerámicas (Carreño, 2005). Actualmente, este compuesto se emplea en la fabricación de cementos de uso odontológico.
La vid silvestre, un taxón amenazado por causa antrópica
La tesis doctoral sobre la situación de la vid silvestre en Europa, llevada a cabo por Claire Arnold de la Universidad de Neuchatel, puso de manifiesto la regresión sufrida por esta vitácea desde el siglo XIX hasta el presente siglo (Arnold, 2002).
Las explotaciones de los bosques de ribera, para uso hortícola y forestal, así como el impacto causado por las especies fúngicas norteamericanas fueron, probablemente, las causas principales de la destrucción de las vides silvestres. A ello, hay que añadir la construcción de embalses y de vías de comunicación y sus tareas de mantenimiento, donde, principalmente, el desbroce indiscriminado de las cunetas constituye un enemigo de las parras (Arnold, 2002) (Figura 12). Respecto a otras obras públicas, en los últimos años se ha producido una drástica pérdida de ejemplares en las playas vizcaínas de Muskiz y Atxabiribil.
Figura 12. Obra pública que muestra la destrucción de los lugares en donde vive la vid silvestre

Fuente: archivo personal.
A comienzos del siglo XIX, en La Algaida (Sanlúcar de Barrameda, Cádiz), actual preparque de Doñana, debía existir una población enorme; así lo refería Rojas Clemente (1807: 105 texto y nota):
Se propagan espontáneamente en este sitio varios vidueños perfectamente caracterizados, encontrándose de cada uno individuos muy viejos, otros recién nacidos y de todas las edades intermedias, pero ninguno que desmienta su casta o afecte a la forma ni demás propiedades que distinguen a las compañeras... En estos mismos sitios forman los vidueños bravios, selvas impenetrables, cenadores magníficos, pabellones graciosísimos, grutas, plazas, caminos cubiertos, veredas tortuosas encrucijadas, laberintos, murallas arcos, columnas y otros mil caprichos originales e indescriptibles.
En 1994 encontramos únicamente 50 ejemplares de dicha población dioica. Un incendio acaecido hace unos 20 años y algunos cortes selectivos de ejemplares de gran tamaño que subían por los pinos han reducido la histórica población a únicamente cuatro ejemplares.
Cuando se destruyen las vides silvestres, también desaparecen varias especies de insectos y ácaros que son enemigos naturales de las plagas, muchas de las cuales no se encuentran en los viñedos por el uso de pesticidas. También se ven afectadas las micorrizas asociadas a las raíces. La máxima biodiversidad de las mismas se ha registrado en la población de la Cascada de Peñaladros (Valle de Angulo, Burgos) (Ocete et al, 2015a), donde muchos de sus ejemplares han sido devastados hace tres años.
Otro problema es que los portainjertos e híbridos productores directos, traídos a Europa desde Norteamérica para paliar la crisis filoxérica, se han propagado indiscriminadamente, convirtiéndose en auténticas plantas invasoras que están desplazando a las vides silvestres euroasiáticas de sus ecosistemas naturales, como puede apreciarase en zonas protegidas como, por ejemplo el Montseny y la Sierra de Cazorla. Este hecho se produce porque las viníferas autóctonas estadounidenses tienen el mismo hábitat que nuestras silvestres (Zecca et al, 2010).
Importancia actual y futura de la vid silvestre como recurso fitogenético
Como se ha indicado anteriormente, las vides silvestres constituyen el parental de las actuales variedades de cultivo. El artículo sobre las poblaciones de vid silvestre en el Valle del Roncal (Navarra) halladas por Martínez de Toda, Llop y Sancha, publicado por Antonio Castillo en el diario El País, el 16 de junio de 1990, dada su importante repercusión mediática, sirvió para catalizar el interés por estas lianas en España.
En el caso de la Península Ibérica, los estudios genéticos han demostrado que más del 75% de las variedades de cultivo tradicionales contienen el clorotipo A, que es el que poseen las poblaciones silvestres de este territorio. Como contrapunto, en la zona de Transcaucasia, la cuna de la Viticultura, sus poblaciones silvestres y variedades cultivadas exhiben los clorotipos C y D (Arroyo-García et al, 2016). Este hecho hace pensar que nuestro territorio pudiera haber sido un centro secundario de domesticación. De todas formas, parece que, al menos, nuestras vides silvestres han ido hibridando por vía materna con las variedades hermafroditas aportadas por fenicios, cartagineses, romanos, árabes, etc.
La variedad femenina Hebén, citada desde el siglo XVI (Alonso de Herrera, 1513) es actualmente vestigial en el viñedo español (Cabello et al, 2011). No obstante, se la considera la progenitora de un amplio conjunto de vidueños ibéricos, como Airén, Mollar Cano, Viura y Xarello (Ghaffari et al, 2014; Martínez-Zapater e Ibáñez, 2015).
Cabe preguntarse entonces ¿qué pueden aportar las vides silvestres a la mejora de variedades de cultivo? Aparte de ciertas consideraciones fitosanitarias, ya comentadas, servirían para la obtención de nuevos portainjertos, siempre hibridadas con especies norteamericanas resistentes, porque, como hemos visto, nuestras vides autóctonas son sensibles a la fase radicícola de la filoxera. No obstante, pueden aportar dos características importantes: resistencia al encharcamiento, dadas las condiciones edáficas de sus hábitats naturales, y a la caliza activa. En efecto, algunas plantas silvestres pueden llegar a tolerar en invernadero hasta un 40% de caliza activa (Cambrollé et al, 2014, 2015).
Desde el punto de vista enológico, hay que resaltar que algunas microvinificaciones con racimos silvestres llegan a tener hasta un 14,5% v/v de etanol. Respecto a los polifenoles totales, la cifra media se sitúa en torno a 80 mg/l, lo que indica que se trata de vinos con una dotación polifenólica elevada, capaces de soportar una crianza prolongada. Además estas muestras de vinos conservan un buen grado de acidez, incluso en clima cálido mediterráneo, con valores de pH comprendidos entre 3,1 y 3,5. También exhiben una importante intensidad de color, que puede superar el valor de 26, dada su vocación tintorera potente por su elevada concentración antociánica, por encima de los 300 mg/l (Meléndez et al, 2016) (Figura 13). Téngase en cuenta que por ejemplo, en Andalucía, para dar color a la gama de vinos tintos actuales, se tiene que recurrir en muchos casos a la variedad foránea Syrah.
Figura 13. Vino procedente de vid silvestre

Fuente: archivo personal.
Estas dos últimas características son de gran importancia de cara a enfrentar el reto del cambio climático, plantando incluso parras femeninas silvestres y obteniendo nuevos cultivares con variedades tradicionales ibéricas y material silvestre.
Por otra parte, actualmente el INCAVI está llevando a cabo el aislamiento y determinación de especies de levaduras silvestres, ya que algunas de las mismas podrían tener aplicación enológica.
Las poblaciones de vid silvestre albergan un importante pool genético, ya que su reproducción sexual conlleva la formación de semillas que dispersan fundamentalmente las aves. De hecho, su diversidad natural constituye la principal fuente de variación genética para las variedades de cultivo, puesto que la vid silvestre ha venido evolucionando libre de la selección artificial humana.
El viñedo, al igual que otros cultivos, se encuentran sometidos a una constante y preocupante erosión genética. Hay que tener en cuenta que el comienzo de la viticultura pasó por una primera selección de ejemplares hermafroditas y determinados procesos de domesticación. Así, se fueron propagando solo aquellas plantas que se ajustaban a la necesidad de aquellas sociedades. Se buscaban ejemplares hermafroditas con alto rendimiento de cosecha, mayor contenido azucarado y mejor resistencia a las condiciones medioambientales de cada zona vitícola, así como una mayor tolerancia/resistencia a especies parásitas (Forni, 2004).
La reconstrucción del viñedo filoxerado supuso una importante pérdida varietal. Por ejemplo, antes de la llegada de esa plaga en 1878, Rojas Clemente (1807) citaba 117 variedades de cultivo en Andalucía. Actualmente, solo los vidueños Pedro Ximénez (Pero Ximen), Zalema y Palomino Fino se encuentran bien representados en el viñedo de esa región. La mayoría de los restantes son meramente vestigiales o han desaparecido.
Por otra parte, la creación de las diversas Denominaciones de Origen limitó el número de variedades admitidas en cada una de ellas. La necesidad de plantar barbados injertados con certificado sanitario también restringe el número de clones ofertados en el mercado.
Si nos fijamos en la variedad Tempranillo de La Rioja, las cepas antiguas producían bayas de tamaño mediano, con un máximo de dos semillas. En los clones actuales, muy productivos, ha aumentado el volumen del grano y el número de pepitas suele oscilar entre tres y cinco. Algunos están ya dando problemas de falta de acidez y de color (comunicación personal de D. Manuel Ruiz Hernández). Además hay que realizar una doble vendimia, donde la primera es tirarla al suelo, con el fin de no sobrepasar el cupo de 6.500 kg/ha, impuesto por el Consejo Regulador de la Denominación de Origen Calificada Rioja.
En general, los efectos del cambio climático obligan a realizar vendimias anticipadas que hacen variar la composición química de las bayas, modificando las características organolépticas de los vinos (Martínez et al, 2016).
Además, asistimos a un empleo creciente de variedades foráneas internacionales, cuyos nombres empezaron a sonar en España a través de telefilmes norteamericanos como los de la serie Falcon Crest, emitida en la década de 1980. El empleo de los cultivares Chardonnay, Sauvignon Blanc, Cabernet Sauvignon, Merlot, Shyrah, etc., tienden a hacer más homogénea la oferta de vinos a nivel mundial. Puede que este sea el espúreo tributo a pagar por causas inherentes al proceso de globalización, que también afecta a nuestras variedades de uva de mesa y de pasificación. Las variedades apirenas importadas cada día ganan más terreno a nuestras variedades tradicionales, como la Ohanes, Aledo, Don Mariano, Moscatel de Alejandría, etc.
De los últimos párrafos, se deduce que se ha reducido drásticamente la base genética sobre la que actúa la selección natural, dada la escasa biodiversidad de las nuevas plantaciones. Este hecho lamentable puede derivar en problemas cualitativos de las cosechas, causados por el cambio climático, como ya se ha referido, y la posible proliferación de nuevas plagas y enfermedades. Estamos asistiendo a un fortísimo aumento de infestación del tornillo, Xylotrechus arvícola (Olivier) (Coleoptera, Cerambycidae); de la araña amarilla común, Tetranychus urticae Koch, del mosquito verde; Jacobiasca lybica (Bergenin & Zanon) (Homoptera: Cicadellidae), etc., así como al incremento imparable de infección por el complejo de hongos de madera, entre otros problemas sanitarios.
La OIV (2010) enfatizaba la necesidad de conservar el germoplasma de vid silvestre. Asimismo, han sido numerosos los trabajos de investigación llevados a cabo dentro del proyecto Europeo COST Action FA 1003 East-West Collaboration for Grapevine Diversity Exploration and Mobilization of Adaptive Traits for Breeding. Como consecuencia de ello, en la asamblea de fin de proyecto, se tomó el acuerdo de elevar una propuesta de protección a la Unión Europea (Ocete et al, 2015b).
Pese a la importancia pasada, presente y futura de este recurso fitogenético que nos ocupa, en el Estado español no existe ninguna figura específica para su protección, como sí ocurre en otros países de la Unión Europea; tal es el caso de Francia, Alemania y Hungría. Varios de los firmantes de este artículo llevan casi una treintena de años intentándolo en diversos organismos de las Administraciones Central y Autonómicas sin ningún éxito. A nuestro entender, esta indiferencia es inexplicable, dada la importancia social y económica de la Viticultura y de los sectores derivados de la misma. España cuenta con unas 954.000 ha de viñedo, la mayor superficie mundial concentrada en un solo país. Urge contar con dicha legislación para impulsar la conservación in situ y ex situ de este patrimonio sobre el que se cimenta nuestra cultura mediterránea y europea de la Viña y el Vino. Además, las rutas con poblaciones de vid silvestre pueden ser un importante atractivo complementario para el creciente sector del turismo enológico. Sería mucho más completo e interesante contemplar los parentales de la variedades cultivadas, visitar los viñedos, bodegas, conocer los métodos de elaboración y catar los caldos producidos en las mismas.
Medidas para intentar paliar la pérdida de este legado silvestre
Bancos de germoplasma
A través de diversos proyectos INIA, se han formado dos bancos de germoplasma. El primero en el actual IFAPA, Rancho de la Merced en Jerez de la Frontera, Cádiz, y el segundo, con un mayor contenido de plantas, en la finca El Encín, perteneciente al IMIDRA, en Alcalá de Henares, Madrid.
En el banco jerezano, una de las acciones llevadas a cabo ha sido la recolección de semillas en la población de La Algaida, con el fin de obtener plántulas para repoblar dicha zona. A nivel provincial, se ha establecido otra colección en la Estación Hortofrutícola de Zalla, dependiente de la Diputación Foral de Vizcaya, otra en la Finca La Orden-Valdesequera, en Badajoz, y una tercera en la Universidad de La Rioja. El IRNAS-CSIC de Sevilla cuenta con un banco de germoplasma in vitro.
Actuaciones in situ
Entre los años 2009 y 2014, se ha procedido a la restauración de las vides en las orillas de la Rivera de Huelva, en las inmediaciones del Pantano de La Minilla (Sevilla), la mayor población encontrada en la zona mediterránea peninsular. Esta zona fue deteriorada por las obras de la autovía de la Ruta de la Plata (Arroyo-García et al., 2016).
En 2014 el Servicio de Biodiversidad de la Diputación Foral de Álava inició la creación de una reserva con plantas autóctonas, entre ellas, vides silvestres de la población del Desfiladero de Sobrón, en los alrededores del Lago de Arreo, un Lugar de Importancia Comunitaria (LIC-ES 2110007) de la Red Natura 2000.
En la actualidad se está desarrollando un proyecto financiado por el Instituto de Estudios Riojanos que implica a la Universidad del País Vasco (Campus de Vitoria/Gasteiz), Universidad de La Rioja y a otros investigadores independientes. Su objetivo principal es el cartografiado de las poblaciones y el recuento exacto de individuos que aún se conservan en dicha región, fundamentalmente, en las cuencas de los ríos Najerilla, Iregua y Rioja.
Actuaciones con posible repercusión económica a corto plazo
En 2008 se puso en marcha un convenio de colaboración entre el IFAPA y la empresa Thalassa Taller de Vino, S.L. para desarrollar el proyecto Caracterización, multiplicación de variedades de vid cultivadas y silvestres autóctonas de Andalucía, con el objeto de contribuir a la modernización del sector vitivinícola de la Serranía de Ronda (Málaga).
En 2012 se estableció un acuerdo de colaboración entre el vivero Fondo Forestal Ibérico y el Laboratorio de Entomología Aplicada de la Universidad de Sevilla para la recuperación de germoplasma silvestre en Andalucía, Castilla y León y País Vasco. Dicho material puede usarse para la restauración de bosques de rivera (Figura 14). Por otra parte, los ejemplares masculinos tienen un alto efecto ornamental en la etapa de floración, donde producen, además, un intenso aroma. Al no producir bayas, se reduce la presencia de abejas y avispas en zonas de jardín muy frecuentes en emparrados tradicionales con plantas hermafroditas. Un ejemplo de uso ornamental puede contemplarse en la zona ajardinada de Bodegas Primicia (La Guardia, Rioja Alavesa).
Figura 14. Plantas de vid silvestre producidas en vivero

Fuente: archivo personal.
En 2012, también se llevó a cabo la recogida de sarmientos de plantas femeninas silvestres para su injerto y plantación en los viñedos de la empresa Término de Miranda, en Miranda de Ebro (Burgos), para realizar las oportunas microvinificaciones, con el fin de su posible aplicación en la elaboración del tradicional chacolí Ojo de gallo.
Conclusión
Es urgente que se tomen medidas para la conservación de las poblaciones de vid silvestre que han sobrevivido hasta nuestros días en España, pese a todas las agresiones sufridas, como bien se explica anteriormente. Se trata de un recurso natural de suma importancia, que en un futuro puede ser clave para mantener el vigor de las vides cultivadas. Sin duda puede contribuir a una mayor diversidad genética de las diferentes variedades de vid que componen los extensos y variados viñedos españoles. Además, a corto y medio plazo, podría tener que utilizarse para obtención de portainjertos, así como para conseguir nuevas variedades de alto valor ecológico, con la posibilidad de mejorar el color de los vinos sin necesidad de la utilización de variedades foráneas. Por ello se insiste que es de suma importancia que se tomen medidas de protección de los ecosistemas tan limitados y desequilibrados en donde aún sobreviven las pocas poblaciones de vid silvestre españolas. La toma de dichas medidas se considera que es urgente, para que surtan vigor a muy corto plazo.
Bibliografía
Alonso, N. “Agricultura, alimentación y entorno vegetal en la Cova de Punta Farisa (Fraga, Huesca) durante el Bronce Medio”. EspailTemps 24 (1995). Quaderns del Departament de Geografia i Historia, Lleida.
Alonso de Herrera, G. Agricultura general. Madrid, Imprenta Real, 1513.
Anzani, R.; Failla, O.; Scienza, A.; De Micheli, L. “Individuazione e conservacione del germoplasma di vite selvatica (Vitis vinifera silvestris) in Italia”. Vignevigne 6 (1993): 5160.
Arnanz, A. M. “Las leguminosas del Cerro de la Cruz (Almedinilla, Córdoba). Presencia de Vicia sativa L. y Vicia ervilia (L.) Willd. en un contexto ibérico”. Complutum 11 (Madrid, 2000): 139-243.
Arnold, C. “Ecologie de la vigne sauvage, Vitis vinifera L. ssp sylvestris (Gmelin) Hegi, dans les forêts alluviales et colluviales d’Europe”. Geobotánica Helvética (2002). Univ. Neuchâtel, Suiza.
Arroyo-García, R. et al. “Multiple origins of cultivated grapevine (Vitis vinifera L. ssp. sativa) based on chloroplast DNA polymorphisms”. Molecular Ecology 15(12) (2006): 3707-3714.
Arroyo-García, R.; Cantos, M.; Lara, M.; López, M.A.; Gallardo, A.; Ocete, C.A.; Pérez, M.A.; Bánati, H.; García, J.L.; Ocete, R. “Characterization of the largest relic Eurasian wild grapevine reservoir in Southern Iberian Peninsula”. Spanish Journal of Agricultural Research 14(3) (2016).
Benito, A.; Muñoz-Organero, G.; De Andrés M.T.; Ocete, R.; García-Muñoz, S.; López, M.A.; Arroyo-García, R.; Cabello, F. “Ex situ ampelographical characterisation of wild Vitis vinifera from fifty-one Spanish populations”. Australian Journal of Grape and Wine Research (2016). Doi: 10.1111/ajgw.
Burjachs, F.; Juliá, R. “Abrupt climatic changes during the last glaciation based on pollen analysis of the Abric Romani, Catalonia, Spain”. Quaternary Research 4 (1994): 308-315. Bustamante, J. La Tierra y los Valles de Mena. Bilbao, Salustegui Editores, 1971.
Buxó, R. Arqueología de las plantas. Barcelona, Crítica/Arqueología, 1997.
Cabello, F.; Ortiz, J.; Muñoz-Organero, G.; Rodríguez-Torres, I.; Benito, A.; Rubio, C.; García-Muñoz, S; Saiz, R. Variedades de vid en España. Madrid, Comunidad de Madrid y Editorial Agrícola, 2011.
Cambrollé, J.; García, J.L.; Figueroa, M.E; Cantos, M. “Physiological responses to soil lime in wild grapevine (Vitis vinifera subsp. sylvestris)”. Environmental and Experimental Botany 105 (2014): 25-31.
Cambrollé, J.; García, J.L.; Ocete, R.; Figueroa, M.E.; Cantos, M. “Evaluating tolerance to calcareous soils in Vitis vinifera subsp. sylvestris”. Plant and Soil 396 (2015): 97-107.
Carbonell, F. Arte de hacer y conservar el vino. Barcelona, Imp. Antonio Brusi, 1820.
Carreño, E. “Las poblaciones de vid silvestre de la Península Ibérica y el origen de las variedades actuales”. Revista Murciana de Antropología 12 (Murcia, 2005): 35-44.
Chamorro, J.G. “Campaña de flotación en el Castillo de Doña Blanca (Puerto de Santa María, Cádiz). Método, muestreo y resultados”. Arqueología medioambiental a través de los macrorrestos vegetales. Madrid, 1991.
Chlilashvili, L. The vine, Wine and Georgians. Authors of the idea. L. Gachechiladze, T. Kandelaki Tbilisi (En georgiano). 2004.
De la Blanchére, H. Les ravageurs des Vergers et des Vignes. París, J. Rothschild Editeur, 1876.
Font Quer, P. Plantas medicinales. ElDioscórides renovado. Barcelona, Labor, 1962.
Forni, G. “Il cammino della vite dall’oriente all’occidente Dall’origine della viticoltura alla sua introduzione in Italia e nel Friuli in particolare. I suoi rapporti con il mondo islamico”. En Del Zan F.; Failla O.; Scienza A. Ampelografia orientale retrospettiva: per una storia del viaggio della vite da oriente a occidente. 2004.
García-Antón, M. Estudio palinológico de los yacimientos mesopleistocenos de Atapuerca (Burgos) reconstrucción paisajística y paleoclimática. Tesis Doctoral. Madrid, Universidad Autónoma, 1989.
Ghaffari, S.; Hasnaoui, N.; Zinelabidine, L.H.; Ferchichi, A.; Martínez-Zapater, J.M.; Ibáñez J. “Genetic diversity and parentage of Tunisian wild and cultivated grapevines (Vitis vinifera L.) as revealed by single nucleotide polymorphism (SNP) markers”. Tree Genetics & Genomes 10(4) (2014): 1103-1112.
Gallardo, A.; Ocete, R.; López, M.A.; Lara, M.; Rivera, D. “Assessment of the Pollen Polymorphism in Populations of Wild Grapevine”. Vitis 48(2) (2009): 59-62.
Guerrero, V. “El palacio-santuario de Cancho Roano (Badajoz) y la comercialización de ánforas indígenas”. Rivista di Studi Finici 19(1) (1995): 49-82.
Iriarte-Chiapusso, M.J.; Gómez-Orellana, L.; Muñoz-Sobrino, C.; Ramil-Rego, P; Arrizabalaga, A. “La dinámica de la vegetación en el NW peninsular durante la transición del Paleolítico medio al Paleolítico superior”. En Montes R.; Lasheras J.A. (eds.). Neandertales cantábricos, estado de la cuestión, Monografías del Museo de Altamira 20 (2005): 231-253, Ministerio de Cultura (Secretaría General Técnica), Madrid.
Iriarte-Chiapusso, M.J.; Muñoz, C.; Gómez-Orellana, L.; Ramil-Rego, P. “Dinámica del paisaje de la Reserva de la Biosfera del Urdabai durante el Holoceno”. Servicio de Publicaciones del Gobierno Vasco. Actas del III Congreso Español de Biogeografia, 2006: 113-117.
Jones, M.; Reed, J. “Agricultural evidence from charred seeds and fruits. The Guadajoz Project. Andalucía in the first millennium a.C”. Vol. 1. Torreparedones and its hinterland. Archaeology Monographs 47 (Oxford, 2000): 403-407.
Laguna, A. Pedacio Dioscórides Anazarbeo, acerca de la materia medicinal y de los venenos mortíferos. Anvers, Juan Latio, 1555.
Lovicu, G.; Farci, M.; Orrú, M.; Ocete, M.E.; López, M.A.; Ocete, R. “Presencia aislada de filoxera y yesca sobre vid silvestre, Vitis vinifera L. subsp. silvestris (Gmelin) Hegi en Cerdeña”. Actas de las XXXI Jornadas de Viticultura y Enología Tierra de Barros. Centro Universitario Santa Ana, Almendralejo, 2009: 105-112.
Maghradze, D.; Salimov, V.; Melyan, G.; Musayev, M.; Ocete, C.A.; Chipashvilli, R.; Failla, O.; Ocete, R. “Sanitary status of the Eurasian wild grapevine in the South Caucasian region”. Vitis 54 (2015): 203-205.
Mai, D.H. “Neue Früchte und Samen aus palaozanen Ablagerungen Mitteleuropas”. Feddes Repert 98 (Berlín, 1987): 197-229.
Martínez, J.L.; Aleixandre Tudó, J.L.; Aleixandre, J.L. “Efectos de los fenómenos producidos por el cambio climático sobre la calidad de los vinos”. Enoviticultura 42 (Barcelona, 2016): 4-26.
Martínez-Zapater, J.M.; Ibáñez, J. “Historia de la vid a la luz de los estudios genéticos”. Historia y arqueología en la cultura del vino. Rafael Francia, coordinador. Instituto de Estudios Riojanos (2015): 15-23.
McGovern, P.E. “Wine and Eurasian grape: Archaeological and Chemical Perspectives on their Origins”. Actas do III Simpósio da Associação International de História e Civilização da Vinha e do Vinho. Funchal, Madeira, 2004: 291-307.
Meléndez, E.; Puras. P.; García, J.L.; Cantos, M.; Gómez-Rodríguez, J.A.; Íñiguez, M.; Rodríguez, A.; Valle, J.M.; Arnold, C.; Ocete, C.A.; Ocete, R. “Evolution of Wild and Feral Vines from the Ega River Gallery Forest (Basque Country and Navarra, Spain) from 1995 to 2015”. Journal international des sciences de la vigne et du vin 50(2) (2016): 65-75.
Negrul, A.M. “Evolution of cultivated forms of grapes”. Comptes Rendus de L’Academie des Sciences de L’URSS 18 (1938): 585-588.
Ocete, R.; López, M.A.; Pérez, M.A.; Del Tío, R.; Lara, M. Las poblaciones españolas de vid silvestre: características de un recurso fitogenético a conservar. Madrid, Monografias INIA/Agrícola 3, 1999.
Ocete, R.; López, M.A.; Gallardo, A.; Pérez, M.A.; Troncoso, A.; Cantos, M.; Liñan, J.; Arnold, C.; Pérez-Camacho, F.; Lara, M. Las poblaciones andaluzas de vid silvestre, Vitis vinifera L., subespecie sylvestris (Gmelin) Hegi: Estudio ecológico, ampelográfico, sanitario y estrategias de conservación. Sevilla, Consejería de Medio Ambiente Junta de Andalucía, 2004.
Ocete, R.; Cantos, M.; López, M.A.; Gallardo, A.; Pérez, M.A.; Troncoso, A.; Lara, M.; Failla, O.; Ferragut, F.J.; Liñán, J. Caracterización y conservación del recurso fitogenético vid silvestre en Andalucía. Sevilla, Consejería de Medio Ambiente, Junta de Andalucía, 2007.
Ocete, R.; Arnold, C.; Failla, O.; Lovicu, G.; Biagini, B.; Imazio, S.; Lara, M.; Maghradze, D.; López, M.A. “Considerations on the European wild grapevine (Vitis vinifera L. subsp. sylvestris (Gmelin) Hegi) and Phylloxera infestation”. Vitis 50(2) (2011a): 97-98.
Ocete, R.; Muñoz-Organero, G.; López, M.A.; Pérez, M.A.; Benito, A.; Cabello, F.; Valle, J.M. “Environmental, sanitary and ampelographic characterization of wild grapevine in western Pyrenees (Spain, France)”. Journal International des Sciences de La Vigne et du Vin 45 (2011b): 1-12.
Ocete, R.; Armendáriz, I.; Cantos, M.D.; Álvarez, M.D.; Azcón, R. “Ecological characterization of wild grapevine habitats focused on arbuscular mycorrhizal symbiosis”. Vitis 54 (2015a): 207-211.
Ocete, R.; Fevereiro, P.; Failla, O. “Proposal for the wild grapevine (Vitis vinifera L. subsp. sylvestris (Gmelin) Hegi) conservation in the European countries”. Vitis 54 (2015b): 281282.
OIV (Organización Internacional de la Viña y el Vino). Descriptor list for grape varieties and Vitis species Paris, A. Dendon, 2009.
-----. Conservación de los recursos genéticos de la vid. Resolución OIV/VITI 424/2010.
Tiblisi, 25 de junio, 2010.
Postigo, J.M.; Burjachs, F.; Gómez-Manzaneque, F.; Morla, C. “A palaeoecological interpretation of the lower-middle Pleistocene Cal Guardiola site (Terrassa, Barcelona, NE Spain) from the comparative study of wood and pollen samples”. Review of Palaeobotany and Palynology 146 (2007): 247-264.
Quer, J.; Gómez Ortega, C. Continuación de la Flora Española ó Historia de las plantas de España 6. Madrid, Joaquín Ibarra Impresor de Cámara de S. M., 1784.
Rivera, D.; Walker, M.J. “A review of paleobotanical findings of early Vitis in the Mediterranean and on the origin of cultivated grape-vines, with special reference to new pointers to prehistoric explotation in the Western Mediterranean”. Review of Palaeobotany and Palynology 6 (1989): 205-237.
Rodríguez-Ariza, M.O.; Ruiz, V.; Ros, M.T. “Paleobotany of a bronce Age community, Castellón Alto (Galera, Granada, Spain)”. Actes du colloque de Périgueux 1995 (1996): 191-196.
Rojas Clemente; Rubio, S. Ensayo sobre las variedades de vid que vegetan en Andalucía. Madrid, Villalpando, 1807.
Rovira Buendía, N. “Semillas y frutos arqueológicos del yacimiento calcolítico de Las Pilas (Mojácar, Almería)”. Complutum 11 (Madrid, 2000): 191-208.
Schumann, F. “Berichte über die Verwendung der Wildrebe Vitis vinifera L. var. silvestris Gmelin”. Die Wein-Wissenschaft 26 (1971): 212-218.
Torres-Vila, L.M.; Mosquera-Müller, J.L. “Caracterización de las pepitas de vid de una ofrenda funeraria romana bajo-imperial (siglos IV-V) de Los Bodegones (Mérida)”. Revista de Arqueología 7 (2001): 455-465.
Vavilov, N.I. Studies on the origin of cultivated Plants. Leningrad, 1926.
Zecca, G.; De Mattia, F.; Lovicu, G.; Labra, M.; Sala, F.; Grassi, F. “Wild grapevine: silvestris, hybrids or cultivars that escaped from vineyards? Molecular evidence in Sardinia”. Plant Biology 12 (2010): 558-562.
Recibido: 3/1/2017 Aprobado: 5/5/2017
Revista RIVAR es editada bajo licencia CREATIVE COMMONS
